Ancient human genomes provide evidence for surprisingly dynamic, interconnected population histories

“Ust’-Ishim” has now joined the ranks of human fossil find sites that biological anthropology students will be conscientiously writing down on index cards, as they prep to memorize spelling and key associated facts, like geographic location and geological age. It thus joins the likes of Hadar, of Dmanisi.
OK. As with those other sites, “Ust’-Ishim” itself should not really be the purpose of any memorization exercise. (Although you can really get into spacing out and repeating “Ust’-Ishim,” as it rolls ticklingly off the tongue … and as your concentration on studying fades … Now, rested and focused from your meditation, back to the important point.)
What is it that’s important to remember here? The site provides a kind of mnemonic tag for learning something new about our part in the natural world. “Ust’-Ishim” is now a heavy-stock, clearly printed and embossed tag for these two facts:
- Anatomically modern humans spread out of Africa and–over many generations of recurring, slight, but significant population growth, over the time period 60,000-30,000 years ago (very roughly)–they decisively made an outsized contribution to ancestry of all later populations outside of Africa, thus shaping worldwide patterns of genetic diversity, right up to the present day.
- But anatomically modern humans (AMH) did not spread into a Eurasian landmass devoid of people. When AMH population growth and expansion into Eurasia began to reach a (quite low but non-zero) escape velocity around 60,000 years ago, at least some AMH groups–with what was then relatively recent African ancestry–mixed and interbred at substantial rates with indigenous Neandertal and other Eurasian populations.
So, the basic recap: major AMH population expansion across Eurasia after 60,000 years ago, and admixture between AMH and indigenous “archaic” human groups.
To be sure, there’s plenty of controversy and debate about why AMH groups successfully and rapidly grew and spread out of Africa when they did … and then persistently pushed the geographic frontiers of the genus Homo, especially northward into arctic latitudes, southeastward into Australia, and eastward into the Americas. Moreover, there’s plenty of disagreement and uncertainty about when the initial, significant pattern of interbreeding occurred, how long it persisted, and–perhaps most significantly–why it was not followed by further interbreeding, but rather population competition and archaic human extinction.
Ust’-Ishim: Tracking Anatomically Modern Humans from Africa to (the Far Reaches of) Eurasia
Now, the precise location of the Ust’-Ishim find site is not easy to get to. At least not anymore. The human femur was discovered in 2008 by a local mammoth ivory carver. (The River Irtysh valley, in western Siberia, has a lot of mammoth and woolly rhino fossils.) Later, after reporting the bone to the authorities, he was not 100% sure where he had actually come upon it. Forensic anthropologist Aleksei Bondarev received the femur and quickly realized that this was hardly the remains of a recently missing person. Instead, it was probably associated with paleontological deposits dating to the last Ice Age. Despite uncertainty about the exact find spot, the surrounding river terrace sediments include not only mammoth and woolly rhino, but also wild horse, European bison, musk ox, and reindeer bones. Moreover, they date roughly to 50-25,000 years ago, at a time when the River Irtysh would have cut through a forest tundra landscape.
A small sample of bone from the femur was sent to the Oxford Radiocarbon Accelerator Unit at Oxford University, and isolation and ultrafiltration of intact bone collagen provided plenty of organic carbon for measuring the 14C level. This radioactive isotope of carbon can give an accurate indication of when the femur was actually part of a living, breathing human body. The 14C date came out to ca. 45,000 years ago (plus or minus a few millennia … ). Measurement on a second sample confirmed the initial radiocarbon result, providing an unusually accurate (and reasonably precise) geological age for the femur.
Now, the two nearly identical radiocarbon dates on collagen directly extracted from the Ust’-Ishim femur are pretty interesting on their own. This is because the femur’s anatomy–with an especially strong pilaster (seen in posterior view on the photo above, posted with Ewen Callaway’s summary article on the Ust’-Ishim research in Nature)–fits an anatomically modern human pattern. Although some western Eurasian human skeletal remains from this general time period (ca. 60-30 thousand years ago) exhibit a mixture of distinctively Neandertal traits along with then-contemporary anatomical features found in Africa and eastern Asia, the Ust’-Ishim femur is morphologically most similar to those of later anatomically modern humans who lived in Europe around 30 thousand years ago, after the virtual disappearance of what had long been regionally dominant patterns of Neandertal anatomy and genetic variation. In other words, Ust’-Ishim may be the oldest known site associated with the decisive post-60,000 BP (before present) population expansion of anatomically modern humans into Eurasia–a demographic expansion thoroughly interconnected with Neandertal extinction.
And where we have collagen very well preserved in ancient bone, it is also possible that DNA from the bone cells might also be trapped, fairly intact, within the bone. This is where the really big scientific news comes in. Indeed, the title of the scientific article in Nature (Fu et al., 2014) is “Genome sequence of a 45,000-year-old modern human from western Siberia.”
The individual represented by this single femur may now be characterized–at least in terms of genetic inheritance–by a carefully reconstructed and aligned genome, including most of the three billion DNA base pairs each human has in the 46 chromosomes and the mitochondrial DNA found in each somatic cell.
Among other things, the reconstructed genome shows the presence of an X and a Y chromosome. This individual is a male.
And consistent with the apparent anatomically modern pattern in the femur’s morphology, this individual is mostly similar–in a comprehensive series of DNA markers that vary in the documented modern human and ancient Neandertal and “Denisovan” genomes–to living humans … and to two recently reconstructed genomes from later Stone Age Eurasian hunter-gatherer skeletons (one from the Upper Paleolithic Mal’ta site in south-central Siberia, dating to ca. 24 thousand years ago, and the other from the Iberian Mesolithic site of La Braña-Arintero, ca. 7 thousand years ago).
As John Hawks points out, in his excellent discussion of the Ust’-Ishim research, the DNA inheritance pattern preserved in the femur documents a predominantly (relatively) recent African ancestry profile–one that we extensively share with the Ust’-Ishim male. But Hawks highlights that Ust’-Ishim also differs to a notable extent from living Europeans. Hawks emphasizes that the Ust’-Ishim genome–when compared with ours and with other recently reconstructed, somewhat less ancient genomes–reveals a surprisingly common, dynamic pattern of regional population turnover in Stone Age prehistory. In this human biogeographic dynamic, we see one population expanding geographically; then, some (or much) interbreeding occurs with local populations; yet, sooner or later, the locals get replaced. And they leave relatively little genetic trace after many generations of recombination and fragmentation of their DNA haplotypes, as more immigrants from the expanding population enter the regional gene pool in successive generations. And this seems to have happened recurrently over the millennia of human prehistory.
So, detailed comparisons of similarities and differences in genetic inheritance markers suggest the following. Ust’-Ishim appears to be part of a population expansion out of Africa and into Eurasia–one contributing to the ancestry of later Upper Paleolithic western Eurasians. Yet, Ust’-Ishim is broadly similar to living Eurasians genetically, not because he is a direct ancestor of those of us with more recent Eurasian ancestry … but rather, because he shared anatomically modern, slightly earlier African ancestors with us. As Hawks notes, the DNA analysis data suggest that most living Eurasians mainly descend from different population lineages, including groups who–like Ust’-Ishim’s lineage–also interbred with Neandertals and contributed to the genetic variation in some later Upper Paleolithic populations … but who ended up expanding and–after many millennia–eventually and largely replacing Ust’-Ishim’s descendants.
Considering what Lazaridis et al. (2014) have recently suggested–that the ancient genomic data support the hypothesis that contemporary European genetic variation was shaped by multiple Stone Age AMH populations–the post-60,000 BP spread of anatomically modern humans from Africa into Eurasia likely involved multiple expanding demographic waves. Each would have reached different areas of Eurasia … and at somewhat different times, involving different patterns and levels of interbreeding with Neandertals, other archaic humans, or already-arrived anatomically modern humans. And only a limited subset of these AMH populations later decisively contributed to more recent prehistoric demographic growth waves, from the later Upper Paleolithic period (ca. 30,000-12,000 BP) to Holocene era emergence and expansion of agriculturally-fed populations.
There’s something here that won’t be lost on those who follow research on Neandertals and their relationship to anatomically modern humans. Hawks points out that Ust’-Ishim’s genetic relationship–or lack of it–to living Eurasians suggests that over the past 100,000 years or so, anatomically modern humans have been involved in multiple episodes of prehistoric population expansion and virtual replacement of local ones … and many of these episodes have involved interaction among multiple AMH groups. For the more recent cases of population turnover, which mainly appear to be associated with the adoption and spread of agricultural technology (Skoglund et al., 2012), the same pattern of population biological turnover involved in Neandertal extinction ended up occurring within what everyone agrees is a single biological species, Homo sapiens.
And just to keep a significant part of our multi-tasking focus on biological complexity and the ambiguity that comes with it, consider this. Related biological patterns–including aspects of descent, as well as adaptation–necessarily exhibit fuzzy boundaries. These are imposed by generation-by-generation evolution in sexually reproducing population networks connected by gene flow … From this perspective, anatomically modern humans today–that is, all 7.2 billion of us–may have come to inherit little or nothing in the way of evolutionarily derived Neandertal phenotypes (Sankararaman et al., 2014), but most living non-Africans do harbor bits of DNA that contain distinctive, selectively neutral mutations that–from roughly 400,000 to 50,000 BP–accumulated over the generations within western Eurasian Neandertal populations. Thus, many anatomically modern humans today have mostly African ancestry, with a small amount–1-2%–of Neandertal genetic background thrown in (Fu et al., 2014).
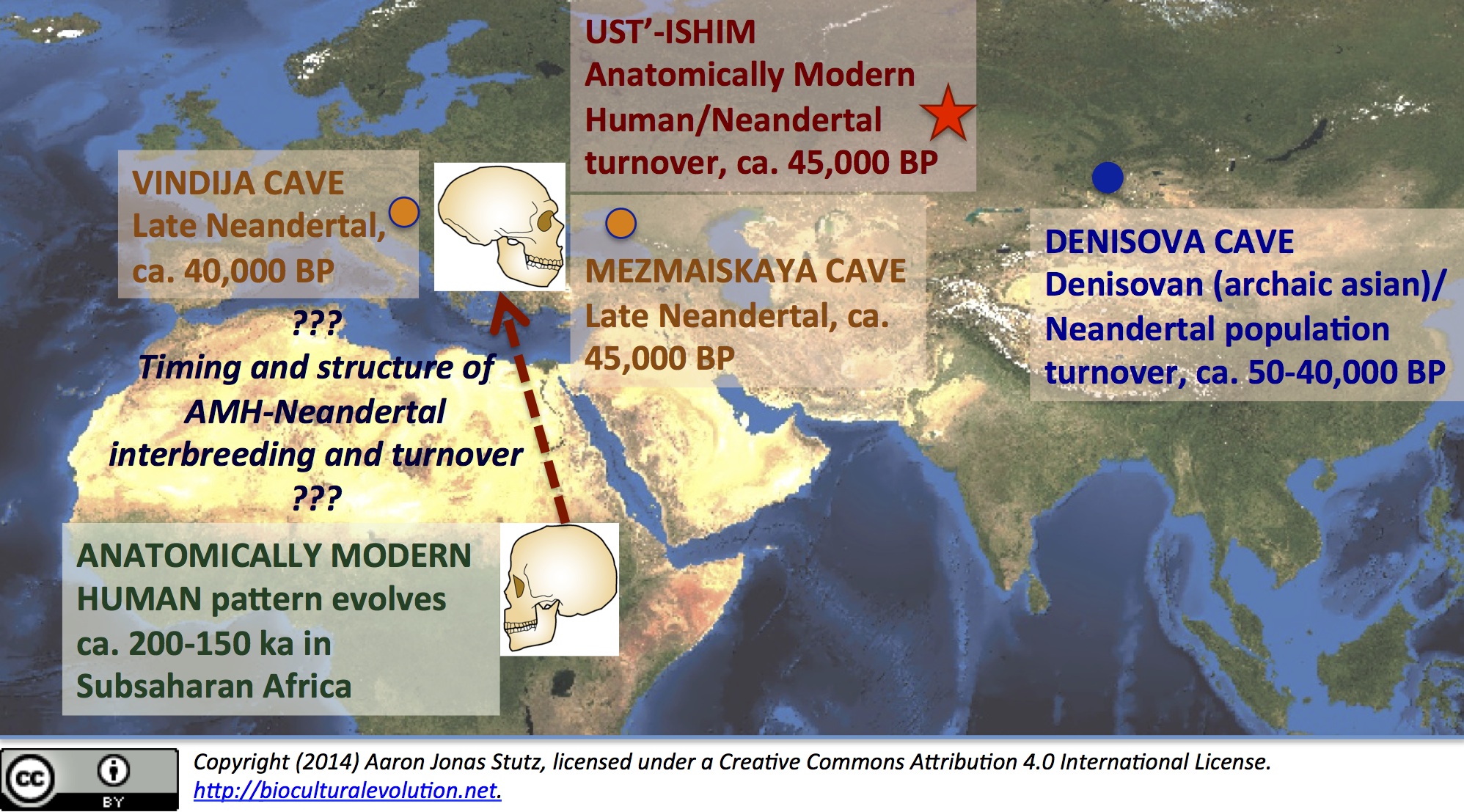
In fact, many people today in Southeast Asia, Melanesia, Polynesia, and aboriginal Australia have a more complex combination of predominantly African background, with small amounts of Neandertal and Asian archaic [so-called “Denisovan”] ancestry (Reich et al., 2010, 2011). Notably, the Ust’-Ishim genome includes no distinctive Denisovan/archaic asian markers. This may be because the Ust’-Ishim male lived and died a few millennia earlier than the only known Denisovan individual–represented by another remarkably well-preserved ancient genome recovered from a humble isolated finger bone (Reich et al., 2010; Prüfer et al., 2013). And speaking of the “Denisovan” genome from Denisova Cave … It is especially relevant that the “Denisovan” finger bone (ca. 40,000 BP) was found stratigraphically overlying a Neandertal toe bone, which dates to ca. 50,000 BP and also yielded whole-genome DNA (Prüfer et al., 2013). Moreover, the “Denisovan”/archaic asian genome preserves evidence of a small amount of Neandertal ancestry. It looks like Denisovan populations also expanded northward, interbreeding with … and then replacing indigenous Neandertals in the Altai Mountains, ca. 40,000 BP. The geographic relationship between Ust’-Ishim and Denisova Cave is shown in the figure below.
What all of this ancient and modern genome data point to is a very dynamic, interconnected population history for the genus Homo. To be sure, researchers have theoretically predicted–and found substantial evidence for–this kind of population turnover with patchy admixture in the spread of prehistoric agricultural and animal domestication technology from the Near East into Europe. The ancient DNA data–even if only obtained from a relatively small number of skeletons, numbering in the hundreds for the mitochondrial DNA only, with still just a few prehistoric remains yielding genome-wide genetic sequences (Olalde et al., 2014)–is beginning to open up a new, remarkable, and increasingly clear window on the surprisingly frequent recurrence of population expansion/interbreeding/turnover dynamics in ancient and recent human evolution. In fact, this complicated population history–exhibiting reticulating connections due to shifting patterns and rates of gene flow–reaches as far back as one milllion years (Prüfer et al., 2013). The new genome-wide results actually support the geneticist Alan Templeton’s (2002) earlier reconstruction of long-term human population history as involving recurrent range expansions against a background of interbreeding, between local groups … and between local and immigrating/expanding groups.
If the evolution of our own genus, Homo, has involved much speciation (that is, evolution of reproductive barriers between what had been inter-fertile populations), then speciation in our genus has been quite different from the evolution of reproductive barriers between very closely related living species, in evolutionary branches nearly as closely related to us. Think chimps and bonobos (Pan troglodytes and Pan paniscus, respectively, with an apparently stable peripatric or allopatric biogeographic distribution and virtually no interbreeding since speciation occurred around 1-2 million years ago [Becquet et al., 2007; Prüfer et al., 2012).
It may be very reasonably argued, then, that what has gone on during most of the evolution of the genus Homo, which has been around perhaps slightly longer than the reproductive barrier between chimps and bonobos in Central Africa, has not involved speciation. Rather, it may be suggested that human evolution over the past 2 million years has involved complex “metapopulation dynamics.” Which means periodic sub-population range expansions and local extinction, recurrent gene flow across sub-population networks, and both local and species-wide adaptation–that is, adaptation through natural selection on variation conserved in the geographically widespread but subdivided population structure (Wolpoff et al., 2000).

At least when it comes to Neandertals, though, there is some ancient DNA evidence for speciation in progress, although neither pre- nor post-mating reproductive barriers seem to have been well-established. Comparisons between ancient DNA and that of living humans with small amounts of Neandertal genetic inheritance suggest that a reproductive barrier between anatomically modern humans and Neandertals may have been evolving, even as Neandertals went extinct. Sankararaman et al. (2014) report genomic evidence for some significant level of reinforcing selection against highly admixed AMH-Neandertal offspring.
It is interesting to note that, if we make the reasonable assumption of persistent millennial-scale weak selection against hybrid offspring, then we must conclude that speciation would not even remotely have been apparent to the members of admixed Stone Age AMH-Neandertal communities, whether from the Near East, Europe, or any other part of western Eurasia. Although opinions will likely continue to diverge on when to divide two interacting, evolving population networks into separate biological species within the genus Homo, it definitely didn’t matter to the people in those populations. And as long as we don’t get too hung up on the often-implicit but scientifically irrelevant symbolism of naming different species, it doesn’t really matter for our understanding of the underlying evolutionary processes. It is more important to figure out why the recurrent-population-expansion/interbreeding/population turnover pattern has so long been characteristic of biocultural evolution in the genus Homo, the recurrently colonizing and interbreeding ape (cf. Wells & Stock, 2008).
REFERENCES CITED
Becquet, C., Patterson, N., Stone, A.C., Przeworski, M., Reich, D., 2007. Genetic Structure of Chimpanzee Populations. PLoS Genet. 3, e66.
Lazaridis, I., Patterson, N., Mittnik, A., Renaud, G., Mallick, S., Kirsanow, K., Sudmant, P.H., Schraiber, J.G., Castellano, S., Lipson, M., Berger, B., Economou, C., Bollongino, R., Fu, Q., Bos, K.I., Nordenfelt, S., Li, H., de Filippo, C., Prüfer, K., Sawyer, S., Posth, C., Haak, W., Hallgren, F., Fornander, E., Rohland, N., Delsate, D., Francken, M., Guinet, J.-M., Wahl, J., Ayodo, G., Babiker, H.A., Bailliet, G., Balanovska, E., Balanovsky, O., Barrantes, R., Bedoya, G., Ben-Ami, H., Bene, J., Berrada, F., Bravi, C.M., Brisighelli, F., Busby, G.B.J., Cali, F., Churnosov, M., Cole, D.E.C., Corach, D., Damba, L., van Driem, G., Dryomov, S., Dugoujon, J.-M., Fedorova, S.A., Gallego Romero, I., Gubina, M., Hammer, M., Henn, B.M., Hervig, T., Hodoglugil, U., Jha, A.R., Karachanak-Yankova, S., Khusainova, R., Khusnutdinova, E., Kittles, R., Kivisild, T., Klitz, W., Kučinskas, V., Kushniarevich, A., Laredj, L., Litvinov, S., Loukidis, T., Mahley, R.W., Melegh, B., Metspalu, E., Molina, J., Mountain, J., Näkkäläjärvi, K., Nesheva, D., Nyambo, T., Osipova, L., Parik, J., Platonov, F., Posukh, O., Romano, V., Rothhammer, F., Rudan, I., Ruizbakiev, R., Sahakyan, H., Sajantila, A., Salas, A., Starikovskaya, E.B., Tarekegn, A., Toncheva, D., Turdikulova, S., Uktveryte, I., Utevska, O., Vasquez, R., Villena, M., Voevoda, M., Winkler, C.A., Yepiskoposyan, L., Zalloua, P., Zemunik, T., Cooper, A., Capelli, C., Thomas, M.G., Ruiz-Linares, A., Tishkoff, S.A., Singh, L., Thangaraj, K., Villems, R., Comas, D., Sukernik, R., Metspalu, M., Meyer, M., Eichler, E.E., Burger, J., Slatkin, M., Pääbo, S., Kelso, J., Reich, D., Krause, J., 2014. Ancient human genomes suggest three ancestral populations for present-day Europeans. Nature. 513, 409–413.
Olalde, I., Allentoft, M.E., Sánchez-Quinto, F., Santpere, G., Chiang, C.W.K., DeGiorgio, M., Prado-Martinez, J., Rodríguez, J.A., Rasmussen, S., Quilez, J., Ramírez, O., Marigorta, U.M., Fernández-Callejo, M., Prada, M.E., Encinas, J.M.V., Nielsen, R., Netea, M.G., Novembre, J., Sturm, R.A., Sabeti, P., Marquès-Bonet, T., Navarro, A., Willerslev, E., Lalueza-Fox, C., 2014. Derived immune and ancestral pigmentation alleles in a 7,000-year-old Mesolithic European. Nature. 507, 225–228.
Prüfer, K., Munch, K., Hellmann, I., Akagi, K., Miller, J.R., Walenz, B., Koren, S., Sutton, G., Kodira, C., Winer, R., Knight, J.R., Mullikin, J.C., Meader, S.J., Ponting, C.P., Lunter, G., Higashino, S., Hobolth, A., Dutheil, J., Karakoç, E., Alkan, C., Sajjadian, S., Catacchio, C.R., Ventura, M., Marques-Bonet, T., Eichler, E.E., André, C., Atencia, R., Mugisha, L., Junhold, J., Patterson, N., Siebauer, M., Good, J.M., Fischer, A., Ptak, S.E., Lachmann, M., Symer, D.E., Mailund, T., Schierup, M.H., Andrés, A.M., Kelso, J., Pääbo, S., 2012. The bonobo genome compared with the chimpanzee and human genomes. Nature.
Prüfer, K., Racimo, F., Patterson, N., Jay, F., Sankararaman, S., Sawyer, S., Heinze, A., Renaud, G., Sudmant, P.H., de Filippo, C., Li, H., Mallick, S., Dannemann, M., Fu, Q., Kircher, M., Kuhlwilm, M., Lachmann, M., Meyer, M., Ongyerth, M., Siebauer, M., Theunert, C., Tandon, A., Moorjani, P., Pickrell, J., Mullikin, J.C., Vohr, S.H., Green, R.E., Hellmann, I., Johnson, P.L.F., Blanche, H., Cann, H., Kitzman, J.O., Shendure, J., Eichler, E.E., Lein, E.S., Bakken, T.E., Golovanova, L.V., Doronichev, V.B., Shunkov, M.V., Derevianko, A.P., Viola, B., Slatkin, M., Reich, D., Kelso, J., Pääbo, S., 2013. The complete genome sequence of a Neanderthal from the Altai Mountains. Nature. advance online publication.
Raghavan, M., Skoglund, P., Graf, K.E., Metspalu, M., Albrechtsen, A., Moltke, I., Rasmussen, S., Stafford Jr, T.W., Orlando, L., Metspalu, E., Karmin, M., Tambets, K., Rootsi, S., Mägi, R., Campos, P.F., Balanovska, E., Balanovsky, O., Khusnutdinova, E., Litvinov, S., Osipova, L.P., Fedorova, S.A., Voevoda, M.I., DeGiorgio, M., Sicheritz-Ponten, T., Brunak, S., Demeshchenko, S., Kivisild, T., Villems, R., Nielsen, R., Jakobsson, M., Willerslev, E., 2014. Upper Palaeolithic Siberian genome reveals dual ancestry of Native Americans. Nature. 505, 87–91.
Reich, D., Green, R.E., Kircher, M., Krause, J., Patterson, N., Durand, E.Y., Viola, B., Briggs, A.W., Stenzel, U., Johnson, P.L.F., Maricic, T., Good, J.M., Marques-Bonet, T., Alkan, C., Fu, Q., Mallick, S., Li, H., Meyer, M., Eichler, E.E., Stoneking, M., Richards, M., Talamo, S., Shunkov, M.V., Derevianko, A.P., Hublin, J.-J., Kelso, J., Slatkin, M., Pääbo, S., 2010. Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature. 468, 1053–1060.
Reich, D., Patterson, N., Kircher, M., Delfin, F., Nandineni, M.R., Pugach, I., Ko, A.M.-S., Ko, Y.-C., Jinam, T.A., Phipps, M.E., Saitou, N., Wollstein, A., Kayser, M., Pääbo, S., Stoneking, M., 2011. Denisova Admixture and the First Modern Human Dispersals into Southeast Asia and Oceania. The American Journal of Human Genetics. 89, 516–528.
Sankararaman, S., Mallick, S., Dannemann, M., Prüfer, K., Kelso, J., Pääbo, S., Patterson, N., Reich, D., 2014. The genomic landscape of Neanderthal ancestry in present-day humans. Nature. 507, 354–357.
Skoglund, P., Malmström, H., Raghavan, M., Storå, J., Hall, P., Willerslev, E., Gilbert, M.T.P., Götherström, A., Jakobsson, M., 2012. Origins and Genetic Legacy of Neolithic Farmers and Hunter-Gatherers in Europe. Science. 336, 466–469.
Templeton, A., 2002. Out of Africa again and again. Nature. 416, 45–51.
Wells, J.C.K., Stock, J.T., 2007. The biology of the colonizing ape. American Journal of Physical Anthropology. 134, 191–222.
Wolpoff, M.H., Hawks, J., Caspari, R., 2000. Multiregional, not multiple origins. American Journal of Physical Anthropology. 112, 129–136.
