Or: It’s All About Evolution, Not Revolution …
by Aaron Jonas Stutz

We’re now paying particular attention to research on the Protoaurignacian, including the notable recent report by Benazzi et al. (2015). We’ll get to their report below. Let’s start with the Protoaurignacian fuss in the first place. There’s pretty good scientific reason for it. Around 2006, Paul Mellars and João Zilhão–two prehistoric archaeologists who otherwise don’t agree about much–separately presented very similar suggestions, which have turned out to be very influential. In a series papers they separately argued that the Protoaurignacian archaeological artifact pattern–or technocomplex, which I will describe in a bit more detail below–has a clear relationship to anatomically modern human expansion into western Eurasia around 40,000 years ago.
Mellars (2006) suggested that the Protoaurignacian artifact pattern is actually a key component of a complex archaeological marker that traces anatomically modern human colonization of western Eurasia … and thus, also of Neandertal extinction.
Zilhão (2006) argued the same thing about anatomically modern humans, but he suggested that anatomically distinctive Neandertals–as opposed to some Neandertal features inherited by some Early Upper Paleolithic people with mainly anatomically modern ancestry–persisted in parts of Europe until ca. 30,000 years ago. On the one hand, there’s no clear threshold of how many unique derived Neandertal traits you need to be “distinctively Neandertal.” On the other, Zilhão and others pointed out that in Iberia, there are some quite distinctively Neandertal fossils, for which a series of radiocarbon dates had placed at roughly 30 kya. The problem now is that those C14 dates were made on incompletely chemically cleaned and filtered collagen-like material surviving in those bones (Wood et al., 2013, 2014). It turns out that all of the Iberian Neandertals so far re-analyzed with the collagen ultrafiltration pretreatment are actually closer to 50-45 kya in age, associated with Middle Paleolithic stone tools.
But what’s important for investigating the Protoaurignacian archaeological phenomenon is that Zilhão’s hypothesis overlaps significantly with Mellars’s. And if the joint Mellars-Zilhão version is right, then there are two very important observations we would expect to be able make … leading to more complete understanding of anatomically modern human / Neandertal biological turnover (a.k.a. Neandertal extinction). First, the oldest Protoaurignacian-like artifact assemblages should be found closer to the African geographic origin of anatomically modern human populations–that is to say, in the Near East. Second, hominin fossils associated with Protoaurignacian archaeological deposits should have anatomically modern–rather than Neandertal–features.
Mellars’s version also predicts two more biocultural evolutionary developments. One is that the chronologically most recent, anatomically identifiable Neandertal fossils should only be found in deposits roughly the same age as Protoaurignacian ones. They should never be substantially younger. His model suggests Neandertals would have undergone a rapid demise due to quite intense ecological competitive exclusion, with repeated waves of AMH demographic growth and dispersal (cf. Mellars & French, 2011). The other is that, in contrast to Zilhão’s formulation–which emphasizes what he saw as fossil anatomical evidence for Neandertal persistence and interbreeding long after the Protoaurignacian technological system was superceded by “classic” Aurignacian systems involving carinated core/scraper and twisted and curved bladelet technologies across much of Europe–Mellars’s model emphasizes that there would have been no significant Neandertal-anatomically modern human population admixture. This is consistent with the view that Neandertals were biologically a distinct species, presumably with a well-defined series of biological barriers to reproduction with anatomically modern humans. As I discuss at the end of this post, the nature of reproductive barriers is a critical question for understanding Neandertal evolution.
Recent research on Protoaurignacian-like sites confirms that–as a technological system–it was widespread across Europe, emerging very early in the Upper Paleolithic period. Moreover, it plausibly developed just as anatomically modern human / Neandertal biological turnover was unfolding.
But do either or both of the joint Mellars-Zilhão predictions about Protoaurignacian origins, spread, and biocultural link to anatomically modern humans actually hold?
Do either or both of the additional Mellars predictions about Neandertal extinction and lack of interbreeding with anatomically modern humans hold?
That’s what this post explores.
Protoaurignacian and Protoaurignacian-like Artifact Assemblages: A Common Technological System, ca. 40 Thousand Years Ago
The most fundamental products of Protoaurignacian technological systems are thin, straight, flat blades (and usually bladelets, formally differentiated from blades in having a length less than 5 centimeters). These sharp-edged stone artifacts are often obtained by striking an already-prepared, narrow-fronted or pyramidal stone core with a “soft” (i.e. not rock) antler, bone or wood hammer. However, such blade-producing techniques occur in systems that other prehistoric societies developed independently in other times and places. The Protoaurignacian technological systems are distinctively recognizable, instead, because they involve production of straight, flat blades and bladelets that are then retouched–which usually involves blunting a sharp edge by removing small, even very very tiny flakes along all or a portion of that edge–in a way that creates instantly recognizable contours.
One key Protoaurignacian retouch pattern is that blades are often finely retouched at the tip distal to end that was initial struck, resulting in a fine, sharp point (see the example above left). Typically, the retouch asymmetrically extends further from the tip on one side of the blade. These artifacts with similar form are known in different parts of western Eurasia as Font-Yves, Krems, or el-Wad points. Moreover, such points would have been effective as projectile spear tips (Shea & Sisk, 2010).
Protoaurignacian systems are often marked by production of straight (or slightly curved) profile bladelets, technically referred to as “Dufour bladelets of the Dufour subtype.” Many of these thin, sharp artifacts are retouched evenly toward the dorsal surface on one side and–inversely–toward the ventral surface on the other. They have been found in the Protoaurignacian-like assemblage from the Early Upper Paleolithic site of Boker A, in the Negev Desert, Israel (Monigal, 2003). They have also been found in the Protoaurignacian site of Trou de la Mère Clochette, in Northeastern France (Szmidt et al., 2010). They have been found in the Early Upper Paleolithic layers from several cave sites in Iran, and in the cave site of Kozarnika in Bulgaria (Olszewski, 1999; Tsanova et al., 2012). Retouched Dufour bladelets may have usually been made in one sitting, intended to function as small serrated saws. Or they may have been resharpened in an arbitrarily consistent way, perhaps first for mounting in a saw or barbed spear, and later refreshed on the opposite, inverse side.
Overall, Protoaurignacian systems strongly contrast with the earlier Paleolithic technological approaches, which emphasize portable multi-use, resharpenable flakes, points, and tools. The Protoaurignacian strategy seemingly aims to produce the maximum amount of cutting edge, with a large number of narrow, thin, replaceable projectile points or knives, from as little flint as possible.
Testing the Mellars-Zilhão Hypothesis about the Protoaurignacian
Joint Prediction 1: Did the Protoaurignacian Originate in the Near East?
Answer: Maybe, but We Just Don’t know Yet
The oldest reliably dated sites with Protoaurignacian technological features are from ca. 42-40 kya (Hershkovitz et al., 2015; Higham et al., 2014; Kadowaki et al., 2015; Stutz et al., in press; Talamo et al., 2012). Currently, measurement uncertainties in the available radiometric dates (variously obtained using C14 and uranium-thorium measurements) make it impossible to discern where Protoaurignacian technologies were first adopted. Thus, we simply do not have a clear enough picture to test the first prediction, one way or another. Did Protoaurignacian-like technological systems first develop with the so-called Initial Upper Paleolithic and Early Ahmarian archaeological technocomplexes in the Levantine zone of the Near East, later spreading with expanding anatomically modern human populations, generation after generation, westward and northward? It’s quite plausible, but we don’t have clear enough “chronometric data,” as we say in the trade.
Wherever the first Protoaurignacian-like system developed locally, the technologies spread relatively quickly throughout much of western Eurasia, likely on the scale of centuries … or at most a couple of millennia. Here, we’re talking something like 50-100 human generations. Although the “Protoaurignacian” archaeological technocomplex was initially named for artifact patterns in key sites from Italy–including one of the main sites studied in the recent report by Benazzi et al. (2015)–the Protoaurignacian in the wide sense may be considered to encompass finds from France in the West to Iran in the East, and central Russia in the north (Bordes, 2006; Hoffecker, 2011; Mellars, 2006; Tsanova et al., 2012; Zilhão, 2011).
Now, there are simply too many alternative ways of systematically knocking blades and flakes off of the same bits of rock, and there are even more ways of retouching or resharpening those blades and flakes into tools with particular piercing, cutting, scraping, or engraving functions. It’s implausible that there were certain environmental conditions occuring around 40 kya that would have necessarily caused the virtually simultaneous adoption of Protoaurignacian technologies, in particular, across a supercontinental area. The Protoaurignacian had to have had a region of origin from which it spread quickly through population dispersal and social interaction. It is indeed remarkable how geographically widespread this particular approach to making stone tools suddenly became, right around or before 40 thousand years ago.
But remember: we don’t know where Protoaurignacian innovations were first developed and adopted. We just know they were widespread, beginning roughly around the same time, 42-40 kya. Although the data aren’t as clear about when Protoaurignacian systems were replaced by other technological approaches, it sems that they did not last too much later than 40 kya in Central and Western Europe (see Banks et al., 2013). However, the Protoaurignacian-like Early Ahmarian system is preserved in the form of artifact assemblages that are likely younger than 35 kya in the Levant (Bar-Yosef & Belfer, Cohen, 2004; Douka et al., 2013; Hershkovitz et al., 2015; Kuhn et al., 2009).
Thus, although Protoaurignacian-like technologies may have persisted in the Levant until as late as ca. 35-30 kya, the Protoaurignacian in Europe was superceded by classic Early Aurignacian technologies roughly around the same time, ca. 38-35 kya, likely over an even wider geographic extent than the Protoaurignacian had reached (Banks et al., 2013; Higham et al., 2011b).
Joint Prediction 2: Are Protoaurignacian Stone Tools Always Archaeologically Associated with Anatomically Modern Human Skeletal Specimens, Never with Anatomically Neandertal Ones?
Answer: Recent Research Suggests So
This is one of the main results of the new report by Benazzi et al. (2015). The authors looked at isolated hominin teeth that were uncovered in layers associated with Protoaurignacian stone tools from two cave sites in Italy, including the large and very rich site of Grotta di Fumane, where long-term excavations have been critical to defining the Protoaurignacian artifact phenomenon itself. Benazzi et al. (2015) have found that the Protoaurignacian-associated teeth are characterized by anatomical traits and mitochondrial DNA fragments statistically linked to modern humans. This is the first time ancient DNA has been recovered from skeletal material apparently associated with Protoaurignacian artifacts. And the hominin mtDNA loci recovered and determined to have been little influenced by contamination exhibit a series of mutations that have been found only in living humans or in ancient DNA from other fossils with anatomically modern features.
The findings presented by Benazzi et al. (2015) are consistent the authors’ previous report that anatomically modern humans are associated with the Uluzzian technocomplex, which is a quite distinct artifact pattern found in many sites in southern Italy (Riel-Salvatore, 2009). The Uluzzian technocomplex likely chronologically overlapped with the Protoaurignacian around 42-40 kya, but it very possibly began earlier. Anatomically modern human skeletal remains have also been found in association with Protoaurignacian-like artifacts (comprising what archaeologists call the Early Ahmarian technocomplex) in Ksar Akil Rockshelter, Lebanon, and Qafzeh Cave, Israel (Bar-Yosef & Belfer-Cohen, 2004; Bergman & Stringer, 1989; Douka et al., 2013).
Of course, any human remains–whether they manifest anatomically modern human or anatomically Neandertal patterns–are rare from the period around 40 kya. The limited available evidence most reasonably supports further scientific inquiry into the joint Mellars-Zilhão hypothesis–that is, that development and adoption of Protoaurignacian technologies across such a wide area in such a brief time has something to do with the emergence of anatomically modern human populations across western Eurasia.
But also of course: as Conard & Bolus (2015) commented on the new results presented by Benazzi et al. (2015), we still don’t fully understand how the Protoaurignacian relates to the establishment of modern anatomically patterns in western Eurasian human populations. Indeed, few paleoanthropologists would agree that we’re basically done. We really need a lot more new fieldwork. And we need laboratory re-analysis of old artifact and skeletal materials.
Mellars Prediction 1: Do the Last Neandertals Go Extinct Around the Same Time as the Protoaurignacian Technocomplex Emerges Across Western Eurasia?
Answer: The Evidence is Pretty Consistent with this Claim …
The previous two sections considered the joint Mellars-Zilhão hypothesis predictions. Support for the general idea is fairly positive. We can’t say for sure that the Protoaurignacian blade and bladelet production techniques, distinctive point-on-blade shapes, and Dufour bladelet retouch patterns were first adopted in the Near East, as anatomically modern human populations were established and then expanded across western Eurasia. The available chronometric analyses—involving mainly radiocarbon dates—just don’t give us enough resolution to rule out that the Protoaurignacian originated in Bulgaria, Italy, France, or even Russia, instead. But where human remains have been identified in stratigraphic association with Protoaurignacian-like artifacts, they have diagnostic modern human anatomy or mtDNA sequences.
As Mellars has argued, the spread of anatomically modern humans out of Africa—and any diagnostic technologies that facilitated their range expansion—constitutes only part of the story. It is vital to remember that anatomically modern humans did not spread out of Africa into a Eurasian landmass devoid of humans. Neandertals and other more ancient Eurasian populations were well established, isolated from Africa by distance—and during cooler, drier periods in the Middle Pleistocene (ca. 800-130 kya), separated by the Saharo-Arabian desert belt. Thus, we finally get back to the issue posed at the beginning of this series of posts. We’re all anatomically modern today, with all or almost all African descent, regardless of where we’re from. So what happened to the humans who had so successfully survived in Eurasia for hundreds of thousands of years? Among the range of hypotheses proposed, Mellars’s own position is that Neandertals did at first survive the Early Upper Paleolithic western Eurasian advance of anatomically modern humans. But, he argues, they were quickly ecologically marginalized, outcompeted by demographically growing, geographically expanding anatomically modern human hunter-gatherer societies (Mellars & French, 2011).
Recent direct, high quality radiocarbon dates on bones with diagnostic Neandertal anatomical patterns support Mellars’s position, at least considered in broad strokes. As we will see below, whether this really constitutes strong support for Mellars’s argument depends on whether the extinction of Neandertal anatomical patterns and most of the derived Neandertal DNA markers also means the extinction of Homo neandertalensis—as Mellars and others would have it. Because it could rather mean an intraspecific macroevolutionary process involving substantial Neandertal-modern human admixture with drift and selection.
We will get to that issue in a moment. First, the rather dirty details of directly (radiocarbon) dating Neandertals.
The figure below summarizes some important information, including:
- the Neandertal skeleton from St. Césaire, France, that has been directly radiocarbon dated to ca. 42-39 kya (Talamo et al., 2012). Although the original site documentation is insufficient to pinpoint the stratigraphic relationship between the unusual Neandertal secondary burial deposit and the surrounding archaeological deposits (Bar-Yosef & Bordes, 2010), the radiocarbon date is consistent with late Chatelperronian age. The so-called Chatelperronian technological system, which is geographically restricted to southwestern France and northern Spain, is subtly but clearly different from the Protoaurignacian (Bordes & Teyssandier, 2011).
- diagnostic Neandertal cranial vault remains from Vindija Cave, Croatia, have been rigorously dated to ca. 38-35 kya, associated with Early Upper Paleolithic stone and bone tools (Higham et al., 2006)
- a Neandertal infant burial from Mezmaiskaya Cave in the northern Caucasus has been carefully dated to ca. 45-42 kya, associated with Late Middle Paleolithic stone tools (Pinhasi et al., 2011).
Continue reading below …
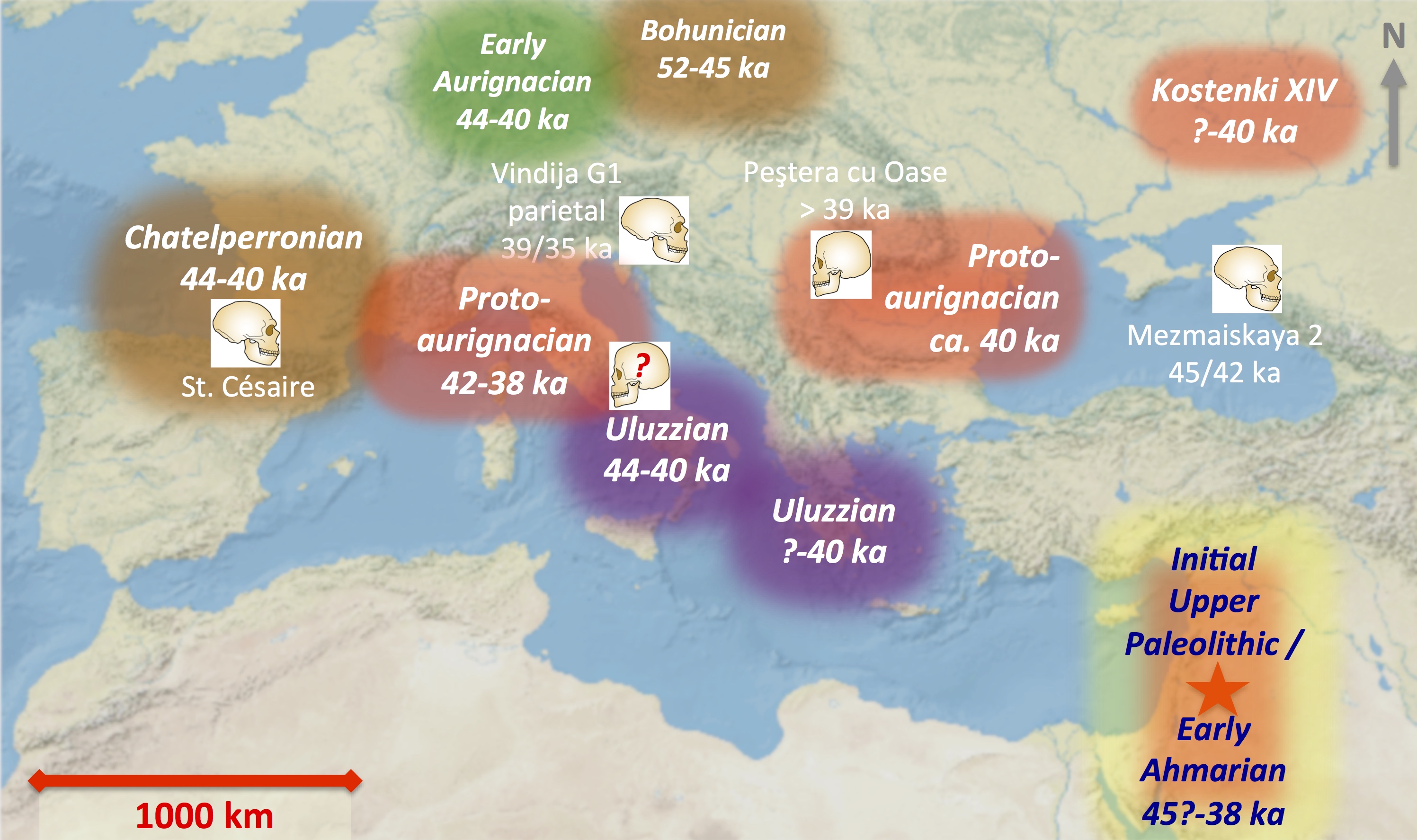
… But the Extinction Process Seems to be Pretty Complex
Now, more equivocal radiocarbon dating results place both anatomically diagnostic Neandertals and modern humans in association with a so-called transitional Middle-Upper Paleolithic system found in northern Europe, known as the Lincombian-Ranisian-Jerzmanowician retouched blade and foliate-shaped biface technocomplex (Flas, 2011), which may date very roughly to ca. 45-40 kya (Higham et al., 2011a; Semal et al., 2009). To be sure, these associations occur at different sites, one in Belgium, the other in England.
In addition, a very interesting, anatomically “ambiguous” jaw bone from Riparo Mezzena in northern Italy is suggestively—but not directly—dated to ca. 40-38 kya, associated with Middle Paleolithic stone tools. The actual radiocarbon date is on an animal bone fragment that is likely stratigraphically associated with the human jaw bone. This mandible is remarkably similar to some very early anatomically modern human specimens (Qafzeh 8, ca. 100 kya in the Levant); roughly contemporary better preserved Neandertals (Spy 1, Belgium, date uncertain, ca. 50-40 kya); and slightly later Upper Paleolithic individuals (Abri Pataud 1, ca. 35 kya). But researchers have recovered recognizably Neandertal mitochondrial DNA from the Mezzena jaw, establishing that it had substantial Neandertal ancestry (Condemi et al., 2013). Although Protoaurignacian layers often overlie Chatelperronian-associated ones at sites in southwestern France, the wider Protoaurignacian technological phenomenon seems to have spread incompletely across the western Eurasian landmass. The evidence reviewed here documents that at least some late Neandertals used other technologies in the ca. 42-38 kya time range, probably including Late Middle Paleolithic and a range of Early Upper Paleolithic ones.
However, in support of Mellars’s position, no anatomically diagnostic Neandertals or any Early Upper Paleolithic humans with predominantly unique derived Neandertal DNA markers likely date to later than 38 kya. The best candidate for later survival of Neandertal traits is the Vindija Cave (Croatia) Vi-208 partial juvenile parietal bone. With some characteristic endocranial Neandertal features (Wolpoff et al., 1981), this cranial vault fragment is carefully and reliably dated to the range 38.5-34.8 kya (95% confidence interval) (Higham et al., 2006). If regional populations with mainly Neandertal anatomy survived past 38 kya, they didn’t survive that much later. Notably, there is no evidence at all of Protoaurignacian artifacts in the Early Upper Paleolithic sequence of Vindija Cave (Ahern et al., 2013).
Mellars Prediction 2: Were the Neandertals a Separate Biological Species of the Genus Homo, having evolved Well-Defined Reproductive Barriers to Anatomically Modern Human Populations?
Answer: Ancient DNA Evidence Now Makes a Compelling Case for Falsifying this Claim
What’s really cool about the recent conference report, presented by Qiaomei Fu (see Callaway, 2015; Gibbons, 2015), is that the quite anatomically modern Oase 1 mandible has not a great-great-great-great-great grandparent’s worth of Neandertal DNA. This is what most humans today have. Rather, the Oase 1 individual (a male, based on identified Y-chromosome markers) has a great-great or great-great-great grandparent’s worth. Now, we’re talking about the kinds of relatively long linked stretches of chromosomes that survive just four or five generations of random recombination. Thus, our first take on this report—presumably soon to be published in much greater detail in a peer-review journal—would be that the Oase 1 individual actually had a really recent ancestor who was also really Neandertal in their own ancestry.
It is important to add that the Oase 1 specimen, recovered as an isolated find in the cave site Peştera cu Oase in Romania, is well-dated—using direct radiocarbon measurement on ultrafiltered collagen fragments and associated uranium-thorium dates on the underlying flowstone in the cave—to between ca. 42-39 kya (Trinkaus et al., 2003). Of course, this means that the Oase 1 mandible is from someone with mixed Neandertal-anatomically modern human ancestry who also happened to live when the Protoaurignacian technological system was spreading, or had spread, across much of western Eurasia. A fragmented skull from another individual was found near the Oase 1 male. Likely of the same age, the Oase 2 skull is also diagnostically anatomically modern human. Yet, like the Oase 1 mandible, it has some isolated traits that are common in Neandertals dating to the previous Middle Paleolithic period (Rougier et al., 2007). Although Peştera cu Oase has no archaeological artifacts associated with the human remains, the nearby site of Romanesti-Dumbrovita has a Protoaurignacian artifact assemblage from a layer dated by thermoluminescence measurements (TL) on heated flint tools to ca. 40 kya (Schmidt et al., 2013).
Basically, the ancient DNA recovered from Oase 1 more strongly confirms what the Mezzena mandible and the varied human remains from the Lincombian-Ranisian-Jerzmanowician sites in Belgium and England suggest. The Early Upper Paleolithic in western Eurasia was the result of social interaction, family formation, and population admixture between anatomically modern humans and Neandertals.
The Take-Home on What We Know about the Protoaurignacian
Although Benazzi et al. (2015) claim that “the Protoaurignacian triggered the demise of Neandertals in this area,” the word “trigger” just doesn’t fit. It’s not consistent with the ancient DNA evidence for occasional, recurrent admixture between Neandertal and anatomically modern human populations. As Qiaomei Fu’s recent published work suggests, admixture likely occurred already among some populations around 55,000 years ago (Fu et al., 2014), although the Neandertal contribution appears to have been diluted by persistent anatomically modern human demographic range expansion out of Africa (see Templeton, 2002). The word “trigger” also suggests a process that is inconsistent with the archaeological evidence. Sure, the Protoaurignacian-like technological systems had a remarkably rapid prehistoric spread and wide geographic footprint. But in the ca. 42-40 kya interval, there were other, very diverse technological strategies for producing, hafting, using, resharpening or discarding stone tools. These strategies geographically surrounded areas of Protoaurignacian adoption. In at least some parts of the Levant, Protoaurignacian-like systems may have just been one of multiple alternatives–all used by the same human population–for producing points, scraping and engraving tools, and quickly disposable flakes and blades (Stutz et al., in press).
To be sure, the work of Mellars and Zilhão has directed our attention to the Protoaurignacian archaeological phenomenon in a very scientifically productive way. Before they began to articulate their arguments, the similarities among Protoaurignacian-like artifact assemblages were not so rigorously documented. And they helped to spur an intense effort to improve the quality of radiometric dating, helping us to clarify the chronology of Protoaurignacian emergence.
So, the most recent–and what I’d argue are quite reliable–results confirm that the Protoaurignacian spread over a remarkable stretch of western Eurasia … and at a critical, albeit long, millennial-scale juncture in anatomically modern human / Neandertal turnover. Still, I’ve summarized available evidence that simply is too imprecise to peg down the geographic source region and spread of the Protoaurignacian to its maximum extent, ca. 40 kya.
It is worth underscoring that the Mellars-Zilhão joint hypothesis–building on Bar-Yosef’s (2000) earlier model of Upper Paleolithic expansion out of the Levant–is logically parsimonious, given broader genetic and fossil evidence for anatomically modern human range expansion out of Africa. We may very well find that the Levant is clearly the source of Protoaurignacian innovations.
In (scientifically, relatively) short, then, the combination of archaeological, ancient DNA, and fossil anatomical evidence suggests that the Protoaurignacian was one of many important biocultural components of a larger, more complex mosaic process of anatomically modern human / Neandertal population biological turnover.
So the Protoaurignacian isn’t the be-all and end-all of Neandertal extinction, in northern Italy, or anywhere else. It didn’t “trigger” anything. One of the most important scientific confirmations that this review emphasizes is that we rarely find a one-to-one correspondence between technological systems and human populations. In the genus Homo–and today, of course, there’s just one quite populous, ecologically important species remaining in the genus–technologies aren’t dependent on the biological or genealogical identities of their makers and users. Populations develop, use, and modify technologies over time, usually for diverse reasons. And populations can change through population booms and crashes, migration, interbreeding, and even large-scale admixture involving ethnogenesis processes. Thus, technological stasis or change might not tell you a clear story about the population change pattern. Although only diagnostically anatomically modern human fossils have been found to associate with Protoaurignacian artifacts, we will also likely find that those Early Upper Paleolithic anatomically modern humans had some ancestors from older, admixed Neandertal-AMH populations.
And if the Neandertals Weren’t Well-Defined as a Separate Species When They Interbred with Anatomically Modern Humans, Then What Were They?
All of this raises an issue we have to focus more on in future work: what were the evolutionary processes at work in Neandertal extinction?
Mellars (2006) may have been right that recognizable Neandertals went extinct by around 38 kya.
But Zilhão (2011) may prove to be right about something else. Building on arguments and ideas developed by Wolpoff and others (see Thorne & Wolpoff, 1992; Wolpoff et al., 2004), he clearly supports a single-species model for anatomically modern human-Neandertal biological turnover. And the recent report of Qiamei Fu’s work, with colleagues, on the ancient DNA from Oase 1 appears to document ongoing interbreeding in the Early Upper Paleolithic period.
The figure below–which I’ve created based on speciation process models summarized by Hvala & Wood (2001)–shows the two most plausible population-interaction scenarios, given the relatively large amount of recurrent admixture with Neandertals during anatomically modern human range expansions out of Africa. What both scenarios share–and what most accounts of speciation in human evolution lack–is an emphasis on ecological reinforcement, involving natural selection for adaptation … and potentially, for reproductive barriers initially shaped by drift in one or both allopatric populations.
In scenario (A), the single-species model emphasizes only differential anatomically modern human demographic contribution–with recurrent range expansions–along with natural selection against Neandertal phenotypes (body proportions? brain organization? skeleto-muscular support for speech communication and facial expression?).
In scenario (B), it is the process of ecological reinforcement that also causes Neandertal extinction, due to ecological competition. Here, Neandertals only come into focus as a well-defined species in the rapid process of their extinction.

References
Ahern, J. C. M., Janković, I., Voisin, J.-L., & Smith, F. H. (2013). Modern Human Origins in Central Europe. In F. H. Smith & J. C. M. Ahern (Eds.), The Origins of Modern Humans: Biology Reconsidered (Second Edition, pp. 151–221). New York: John Wiley & Sons.
Banks, W. E., d’ Errico, F., & Zilhão, J. (2013). Human–climate interaction during the Early Upper Paleolithic: testing the hypothesis of an adaptive shift between the Proto-Aurignacian and the Early Aurignacian. Journal of Human Evolution, 64(1), 39–55. http://doi.org/10.1016/j.jhevol.2012.10.001
Bar-Yosef, O. (2000). The impact of radiocarbon dating on old world archaeology : Past achievements and future expectations. Radiocarbon, 42(1), 23–39.
Bar-Yosef, O., & Belfer-Cohen, A. (2004). The Qafzeh Upper Paleolithic Assemblages: 70 Years Later. Eurasian Prehistory, 2(1), 145–180.
Bar-Yosef, O., & Belfer-Cohen, A. (2010). The Levantine Upper Palaeolithic and Epipaleolithic. In E. A. A. Garcia (Ed.), South-Eastern Mediterranean Peoples Between 130,000 and 10,000 Years Ago (pp. 144–167). Oxford: Oxbow.
Bar-Yosef, O., & Bordes, J.-G. (2010). Who were the makers of the Châtelperronian culture? Journal of Human Evolution, 59(5), 586–593. http://doi.org/10.1016/j.jhevol.2010.06.009
Bar-Yosef, O., & Zilhão, J. (2006). Towards a Definition of the Aurignacian: Proceedings of the Symposium Held in Lisbon, Portugal, [Centro Cultural de Belém], June 25-30, 2002. Instituto Português de Arqueologia.
Benazzi, S., Slon, V., Talamo, S., Negrino, F., Peresani, M., Bailey, S. E., … Hublin, J.-J. (2015). The makers of the Protoaurignacian and implications for Neandertal extinction. Science, aaa2773. http://doi.org/10.1126/science.aaa2773
Bergman, C., & Stringer, C. (1989). Fifty years after: Egbert, an early Upper Palaeolithic juvenile from Ksar Akil, Lebanon. Paléorient, 15(2), 99–111. http://doi.org/10.3406/paleo.1989.4512
Bordes, J.-G., & Teyssandier, N. (2011). The Upper Paleolithic nature of the Châtelperronian in South-Western France: Archeostratigraphic and lithic evidence. Quaternary International, 246(1–2), 382–388. http://doi.org/10.1016/j.quaint.2011.08.001
Callaway, E. (2015). Early European may have had Neanderthal great-great-grandparent. Nature. http://doi.org/10.1038/nature.2015.17534
Conard, N. J., & Bolus, M. (2015). Chronicling modern human’s arrival in Europe. Science, 348(6236), 754–756. http://doi.org/10.1126/science.aab0234
Condemi, S., Mounier, A., Giunti, P., Lari, M., Caramelli, D., & Longo, L. (2013). Possible Interbreeding in Late Italian Neanderthals? New Data from the Mezzena Jaw (Monti Lessini, Verona, Italy). PLoS ONE, 8(3), e59781. http://doi.org/10.1371/journal.pone.0059781
Douka, K., Bergman, C. A., Hedges, R. E. M., Wesselingh, F. P., & Higham, T. F. G. (2013). Chronology of Ksar Akil (Lebanon) and Implications for the Colonization of Europe by Anatomically Modern Humans. PLoS ONE, 8(9), e72931. http://doi.org/10.1371/journal.pone.0072931
Flas, D. (2011). The Middle to Upper Paleolithic transition in Northern Europe: the Lincombian-Ranisian-Jerzmanowician and the issue of acculturation of the last Neanderthals. World Archaeology, 43(4), 605–627. http://doi.org/10.1080/00438243.2011.624725
Fu, Q., Li, H., Moorjani, P., Jay, F., Slepchenko, S. M., Bondarev, A. A., … Pääbo, S. (2014). Genome sequence of a 45,000-year-old modern human from western Siberia. Nature, 514(7523), 445–449. http://doi.org/10.1038/nature13810
Gibbons, A. (2015). Ancient DNA pinpoints Paleolithic liaison in Europe. Science, 348(6237), 847–847. http://doi.org/10.1126/science.348.6237.847
Hershkovitz, I., Marder, O., Ayalon, A., Bar-Matthews, M., Yasur, G., Boaretto, E., … Barzilai, O. (2015). Levantine cranium from Manot Cave (Israel) foreshadows the first European modern humans. Nature, 520(7546), 216–219. http://doi.org/10.1038/nature14134
Higham, T., Compton, T., Stringer, C., Jacobi, R., Shapiro, B., Trinkaus, E., … Fagan, M. (2011a). The earliest evidence for anatomically modern humans in northwestern Europe. Nature, 479(7374), 521–524. http://doi.org/10.1038/nature10484
Higham, T., Douka, K., Wood, R., Ramsey, C. B., Brock, F., Basell, L., … Jacobi, R. (2014). The timing and spatiotemporal patterning of Neanderthal disappearance. Nature, 512(7514), 306–309. http://doi.org/10.1038/nature13621
Higham, T., Jacobi, R., Basell, L., Ramsey, C. B., Chiotti, L., & Nespoulet, R. (2011b). Precision dating of the Palaeolithic: A new radiocarbon chronology for the Abri Pataud (France), a key Aurignacian sequence. Journal of Human Evolution, 61(5), 549–563. http://doi.org/10.1016/j.jhevol.2011.06.005
Higham, T., Ramsey, C. B., Karavanić, I., Smith, F. H., & Trinkaus, E. (2006). Revised direct radiocarbon dating of the Vindija G1 Upper Paleolithic Neandertals. Proceedings of the National Academy of Sciences of the United States of America, 103(3), 553–557. http://doi.org/10.1073/pnas.0510005103
Hoffecker, J. F. (2011). The early upper Paleolithic of eastern Europe reconsidered. Evolutionary Anthropology: Issues, News, and Reviews, 20(1), 24–39. http://doi.org/10.1002/evan.20284
Hvala, J. A., & Wood, T. E. (2001). Speciation: Introduction. In eLS. John Wiley & Sons, Ltd. Retrieved from http://onlinelibrary.wiley.com.proxy.library.emory.edu/doi/10.1002/9780470015902.a0001709.pub3/abstract
Kadowaki, S., Omori, T., & Nishiaki, Y. (2015). Variability in Early Ahmarian lithic technology and its implications for the model of a Levantine origin of the Protoaurignacian. Journal of Human Evolution, 82, 67–87. http://doi.org/10.1016/j.jhevol.2015.02.017
Kuhn, S. L., Stiner, M. C., Güleç, E., Özer, I., Yılmaz, H., Baykara, I., … Suata-Alpaslan, F. (2009). The early Upper Paleolithic occupations at Üçağızlı Cave (Hatay, Turkey). Journal of Human Evolution, 56(2), 87–113. http://doi.org/10.1016/j.jhevol.2008.07.014
Kuhn, S. L., & Zwyns, N. (2014). Rethinking the initial Upper Paleolithic. Quaternary International, 347, 29–38. http://doi.org/10.1016/j.quaint.2014.05.040
Mellars, P. (2006). Archeology and the dispersal of modern humans in Europe: Deconstructing the “Aurignacian.” Evolutionary Anthropology: Issues, News, and Reviews, 15(5), 167–182. http://doi.org/10.1002/evan.20103
Mellars, P., & French, J. C. (2011). Tenfold Population Increase in Western Europe at the Neandertal–to–Modern Human Transition. Science, 333(6042), 623–627. http://doi.org/10.1126/science.1206930
Monigal, K. (2003). Monigal, K. (2003). Technology, economy, and mobility at the beginning of the Levantine Upper Paleolithic. In Goring-Morris & Belfer-Cohen 2003 (pp. 118-133). In A. N. Goring-Morris & A. Belfer-Cohen (Eds.), More Than Meets the Eye: Studies on Upper Palaeolithic Diversity in the Near East (pp. 118–133). Oxford: Oxbow Books.
Olszewski, D. I. (1999). The Early Upper Palaeolithic in the Zagros Mountains. In W. Davies & R. Charles (Eds.), Dorothy Garrod and the Progress of the Palaeolithic: Studies in the Prehistoric Archaeology of the Near East and Europe (pp. 167–180). Oxford: Oxbow Books.
Pinhasi, R., Higham, T. F. G., Golovanova, L. V., & Doronichev, V. B. (2011). Revised age of late Neanderthal occupation and the end of the Middle Paleolithic in the northern Caucasus. Proceedings of the National Academy of Sciences, 108(21), 8611–8616. http://doi.org/10.1073/pnas.1018938108
Riel-Salvatore, J. (2009). What Is a “Transitional” Industry? The Uluzzian of Southern Italy as a Case Study. In M. Camps & P. Chauhan (Eds.), Sourcebook of Paleolithic Transitions (pp. 377–396). Springer New York. Retrieved from http://link.springer.com/chapter/10.1007/978-0-387-76487-0_25
Rougier, H., Milota, Ş., Rodrigo, R., Gherase, M., Sarcinǎ, L., Moldovan, O., … Trinkaus, E. (2007). Peştera cu Oase 2 and the cranial morphology of early modern Europeans. Proceedings of the National Academy of Sciences, 104(4), 1165–1170. http://doi.org/10.1073/pnas.0610538104
Schmidt, C., Sitlivy, V., Anghelinu, M., Chabai, V., Kels, H., Uthmeier, T., … Radtke, U. (2013). First chronometric dates (TL and OSL) for the Aurignacian open-air site of Româneşti-Dumbrăviţa I, Romania. Journal of Archaeological Science, 40(10), 3740–3753. http://doi.org/10.1016/j.jas.2013.04.003
Semal, P., Rougier, H., Crevecoeur, I., Jungels, C., Flas, D., Hauzeur, A., … van der Plicht, J. (2009). New data on the late Neandertals: Direct dating of the Belgian Spy fossils. American Journal of Physical Anthropology, 138(4), 421–428. http://doi.org/10.1002/ajpa.20954
Shea, J. J., & Sisk, M. L. (2010). Complex Projectile Technology and Homo sapiens Dispersal into Western Eurasia. PaleoAnthropology, 2010, 100–122. http://doi.org/10.4207/PA.2010.ART36
Stutz, A. J., Shea, J. J., Rech, J. A., Pigati, J. S., Wilson, J., Belmaker, M., … Nilsson Stutz, L. (n.d.). Early Upper Paleolithic chronology in the Levant: new ABOx-SC accelerator mass spectrometry results from the Mughr el-Hamamah Site, Jordan. Journal of Human Evolution. http://doi.org/10.1016/j.jhevol.2015.04.008
Szmidt, C. C., Normand, C., Burr, G. S., Hodgins, G. W. L., & LaMotta, S. (2010). AMS 14C dating the Protoaurignacian/Early Aurignacian of Isturitz, France. Implications for Neanderthal–modern human interaction and the timing of technical and cultural innovations in Europe. Journal of Archaeological Science, 37(4), 758–768. http://doi.org/10.1016/j.jas.2009.11.006
Talamo, S., Soressi, M., Roussel, M., Richards, M., & Hublin, J.-J. (2012). A radiocarbon chronology for the complete Middle to Upper Palaeolithic transitional sequence of Les Cottés (France). Journal of Archaeological Science, 39(1), 175–183. http://doi.org/10.1016/j.jas.2011.09.019
Templeton, A. (2002). Out of Africa again and again. Nature, 416(6876), 45–51. http://doi.org/10.1038/416045a
Teyssandier, N. (2006). Questioning the first Aurignacian: mono or multi cultural phenomenon during the formation of the Upper Paleolithic in Central Europe and the Balkans. Anthropologie Brno, XLIV(1), 9–29.
Thorne, A. G., & Wolpoff, M. H. (1992). The Multiregional Evolution of Humans. Scientific American, (266(4)), 76–83. http://doi.org/10.1038/scientificamerican0492-76
Trinkaus, E., Moldovan, O., Milota, Ş., Bîlgăr, A., Sarcina, L., Athreya, S., … Plicht, J. van der. (2003). An early modern human from the Peştera cu Oase, Romania. Proceedings of the National Academy of Sciences, 100(20), 11231–11236. http://doi.org/10.1073/pnas.2035108100
Tsanova, T., Zwyns, N., Eizenberg, L., Teyssandier, N., Le Brun-Ricalens, F., & Otte, M. (2012). Le plus petit dénominateur commun : réflexion sur la variabilité des ensembles lamellaires du Paléolithique supérieur ancien d’Eurasie. Un bilan autour des exemples de Kozarnika (Est des Balkans) et Yafteh (Zagros central). L’Anthropologie, 116(4), 469–509. http://doi.org/10.1016/j.anthro.2011.10.005
Wolpoff, M. H., Smith, F. H., Malez, M., Radovčić, J., & Rukavina, D. (1981). Upper pleistocene human remains from Vindija cave, Croatia, Yugoslavia. American Journal of Physical Anthropology, 54(4), 499–545. http://doi.org/10.1002/ajpa.1330540407
Wolpoff, M., Mannheim, B., Mann, A., Hawks, J., Caspari, R., Rosenberg, K., … Clark, G. (2004). Why not the Neandertals? World Archaeology, 36(4), 527–546. http://doi.org/10.1080/0043824042000303700
Wood, R. E., Arrizabalaga, A., Camps, M., Fallon, S., Iriarte-Chiapusso, M.-J., Jones, R., … Higham, T. F. G. (2014). The chronology of the earliest Upper Palaeolithic in northern Iberia: New insights from L’Arbreda, Labeko Koba and La Viña. Journal of Human Evolution, 69, 91–109. http://doi.org/10.1016/j.jhevol.2013.12.017
Wood, R. E., Barroso-Ruíz, C., Caparrós, M., Pardo, J. F. J., Santos, B. G., & Higham, T. F. G. (2013). Radiocarbon dating casts doubt on the late chronology of the Middle to Upper Palaeolithic transition in southern Iberia. Proceedings of the National Academy of Sciences, 110(8), 2781–2786. http://doi.org/10.1073/pnas.1207656110
Zilhão, J. (2006). Neandertals and moderns mixed, and it matters. Evolutionary Anthropology: Issues, News, and Reviews, 15(5), 183–195. http://doi.org/10.1002/evan.20110
Zilhão, J. (2011). Aliens from Outer Time? Why the “Human Revolution” Is Wrong, and Where Do We Go from Here? In S. Condemi & G.-C. Weniger (Eds.), Continuity and Discontinuity in the Peopling of Europe (pp. 331–366). Springer Netherlands. Retrieved from http://link.springer.com/chapter/10.1007/978-94-007-0492-3_25
