Even before Darwin, it should be remembered, scientists contemplated the notion that different kinds of organisms evolve–that is, have evolved in the past and conceivably continue to evolve. And that the evolution of living organisms has to do with adaptation over many generations of reproduction.
The evidence for adaptation through differential reproduction over the generations–driven by differential fitness of the variety of heritable traits in a population … of bacteria, oak trees, people …–is unambiguous. Yet, the implications for how we think about life remain challenging. In my experience, this is particularly the case on two levels. First, biology students have difficulty–especially initially–staying focused on evolution as a gradual process of complex change in differential reproduction over thousands generations. Second, instructors–especially those who have limited background in biology, say, in teaching an introductory anthropology course–have difficulty, more often than one would expect, in combining clear presentation with effective exercises and discussions that would better help students really own and use accurate knowledge of long-term evolutionary processes and outcomes. The evolutionary process is indeed complex, because what changes (again, over many many generations) is simultaneously:
- random mutation rarely but persistently altering how bits of DNA function, sometimes increasing the fitness of a given bit in the environment, sometimes decreasing it
- the characteristics that those more fit bits of DNA build or influence, as they chemically shape phenotypic “vehicles” for their own survival and reproduction (Dawkins 1982a, 1982b)
- variation in adjoining bits of “neutrally fit” DNA that have succeeded in hanging on for the ride while accumulating harmless mutations
- the sustained or accumulated, often ecologically structuring impact of DNA’s chemical, usually complex and indirect influence on those phenotypic vehicles that have already been favored by previous generations of natural selection … with the critical, pervasive effect that DNA indirectly but systemically, cumulatively changes its environment over long time-frames (Odling-Smee et al. 2003)
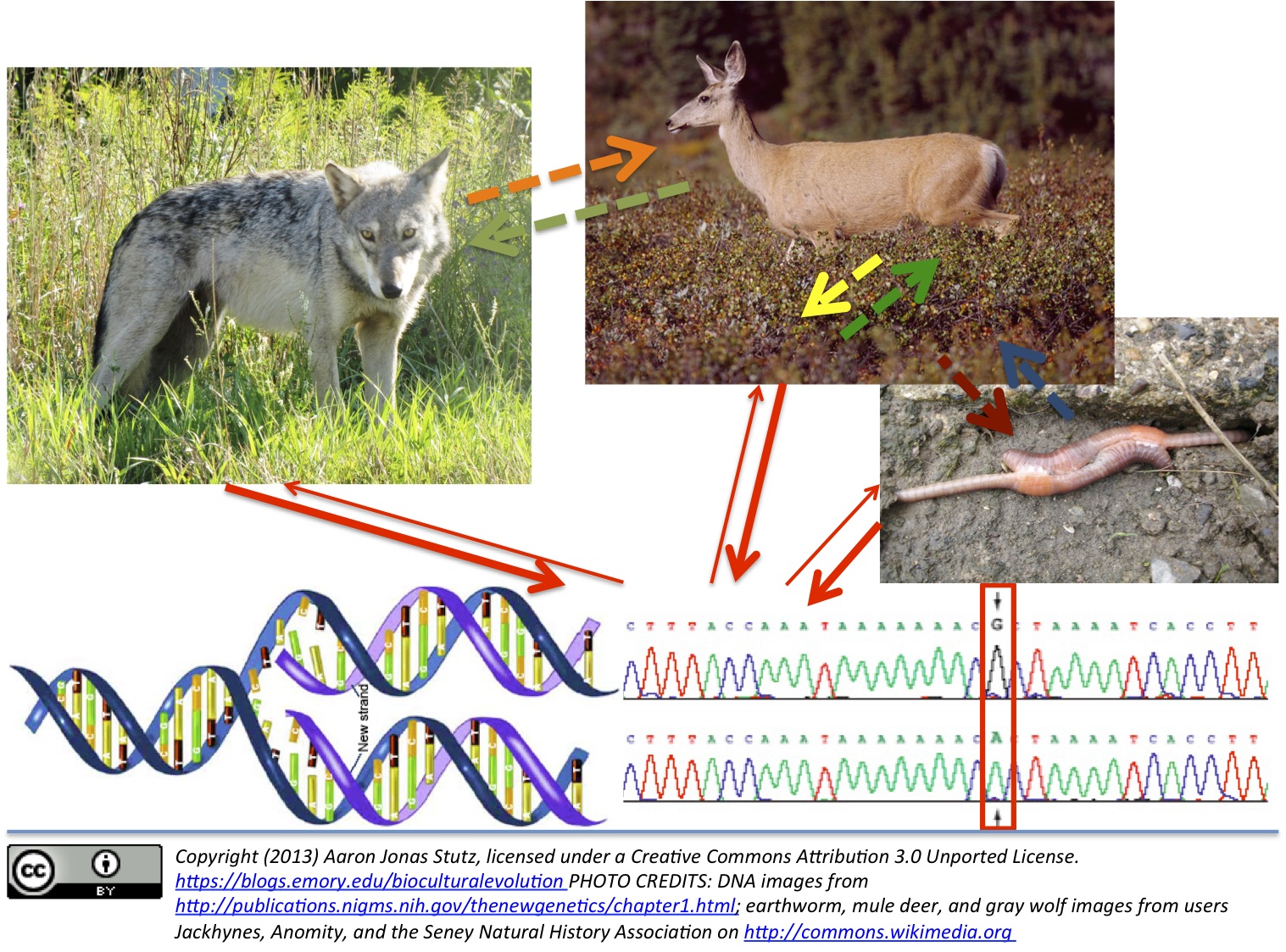
The intricacy of this process may best be conceptualized with a series of abstract notions, like populations and allele frequency change. But at the intro level, just trying to get your head around these thoughts can deaden what provokes curiosity about evolution: birth, maturation, aggression, cooperation, sex, and death in nature. BTW, I had to go with the language “more fit bits” once it popped into my head, since it sounds somewhat disgusting, even a bit obscene. BUT … but hopefully you’ll remember the bigger point about why students find it especially difficult to conceptualize the evolutionary process.
Now note that the complexity of evolution is not irreducible. It is reducible to how DNA chemically functions, impacting its environment, and in turn influencing which DNA-chain variants survive and copy themselves more effectively in that environment. While scientific knowledge always philosophically involves doubt and acknowledgment that there are unknown phenomena in the universe, science is also an active stance for seeking explanations for observable phenomena that involve natural causes or processes. (This perspective is, of course, contra what proponents of “intelligent design” try to sell us on, where it is claimed or hoped that God comes in, every once in a while–allowing us to witness biochemical systems or anatomical structures, at least in some organisms, that exhibit such complexity so as to defy the very physics or chemistry according to which those organisms’ genes are naturally supposed to have evolved or operate. Such claims very simply amount to hoping–because of an a priori belief in divine intervention in nature–that repeatable, independently verifiable observations do indeed record phenomena that scientific inquiry will never ever be able to explain. Among other profound philosophical problems with Intelligent Design–the perspective of which takes a seemingly tactical step away from the Biblical literalist point of departure of so-called “Creation Science”–is this: it is really arbitrary which poorly understood observable phenomenon is left open to scientific curiosity and investigation and which is asserted to be so complex that we can decide here and now that said complexity is evidence of an active divine hand in nature. And this makes Intelligent Design not only a willfully ignorant stance, but also very corruptible, highly susceptible to arbitrary appeals to phony scientific authority.)
Getting back to what evolves in evolution, I would emphasize four absolutely key take-homes from the above brief summary of the evolutionary process:
- Evolution does indeed happen in populations of organisms whenever there is change in DNA diversity from one generation to the next … and thus, evolution is not really that interesting in the short-term … but it is happening.
- IT’S FEEDBACK, PEOPLE: evolution is understood to occur without irreducible complexity, exactly BECAUSE DNA impacts–and thus changes–its own environment, even as random mutation in this theoretically infinitely long biopolymer causes variation in DNA’s very function … including complex interactions among its own chemical products in and around cells … In other words, DNA regulates its own chemical self-copying function–albeit with occasional copying errors (a.k.a. random mutations)–at the same time that it influences other chemical products in its own environment, in which the chemical replication proceeds … so that sometimes DNA has a strong non-nested hierarchical filtering effect on the environment, although most of the time, the environment has a stronger non-nested hierarchical filtering effect on replicating DNA.
- Evolution is interesting in the long-term–that is, over thousands or millions of years–because that’s when all the drama of birth, aggression, cooperation, sex and death emerges. The challenge is that it comes from the complex, occasionally shifting non-nested hierarchical feedback between DNA and its environment, and that leads to much more than anthropomorphically titillating drama … evolution has produced spectacular ecological phenomena that have given the planet Earth a richly dynamic but resilient biosphere–one that’s survived all forms of physical change on the planetary level (plate tectonics and volcanic eruption) and solar system level (oscillations in the Earth’s orbit and variations in solar radiation intensity) over a couple of billions of years.
- It’s worth being interested in points 1. and 2. BECAUSE if you’re interested in 3., then the first two points help you answer a lot of questions and even ask some new, really smart ones.
This is more than just my two cents, as a college instructor in biological anthropology, for why deeper understanding of evolution is worth the effort and how students might begin to answer AND ask better questions about why life works the way it does. As virtually all biologists would emphasize, everyone agrees that evolution in DNA allele frequencies goes on all the time–and in all forms of reproducing populations, from bacteria to trees to whales. But not everyone agrees on the reasons why long-term evolution is most interesting. In scientific discussion and research there arises all kinds of logically derived, yet diverging views about how to analyze the relationship between boring, short-term allele-frequency changes over a few generations and complex, interesting patterns, which may range from DNA function to organismal development to food webs to major adaptive radiations of species … and even to mass extinctions. Among the best known scientific debates about emergent evolutionary process unfolded in the 1970’s and 1980’s: Does speciation, extinction, and adaptation follow a gradual or a punctuated equilibrium proces (Eldredge and Gould 1972; Gingerich 1984, 1985; Gould and Eldredge 1977)? Are common phenotypic traits in populations best assumed to be adaptations shaped by natural selection or structural connectors or place-holders that really have nothing to do with adaptation (Gould and Lewontin 1979; Mayr 1983)? Have most fixed alleles in populations evolved by random drift, despite being selectively neutral (that is, having no influence whatsoever on vehicles that might favor or disfavor replication in the prevailing environment) (Kimura 1983)? Is the branching history of species best reconstructed by phenetic or cladistic analyses of their traits (Gingerich 1985; Stuessy 1987)? Should species be considered individuals (Vrba 1984; Vrba and Eldredge 1984; Vrba and Gould 1986) or should we really mainly focus on the “long reach of the (selfish) gene” (Dawkins 1982b)? No one said that science is easy, but it should be done right, and these theoretical and technical debates were part of the scientific process of trying to get the answer right: asking the right questions, using the right models, making the right measurements, carrying out the right analyses, so that observations could be replicated by other scientists and the research questions, methods, and conclusions all logically connected. Many of these debates have been eclipsed by the flood of data that has come over the past 25 years or so. Now, thanks to increasing computational power and DNA sequencing technologies, biological researchers have begun developing methods that integrate genetic, ecological, biochemical, physiological, anatomical, and behavioral information. And this helps us better understand the connections between areas of biological inquiry that have been all too often over-specialized.
So … despite lingering and often intellectually challenging and productive debates about philosophical approaches to the complex process of evolution, we have to stay focused on key ideas. That is, clear ideas that facilitate our integrating different questions about how DNA-environment feedback–fundamentally involving DNA replication–structures the big picture emergent phenomena. And putting aside debates about whether species are individuals or genes should demand more of our scientific concern, I’d emphasize one clear idea that helps us to getting back to basics, about how DNA-environment feedback can shape diversity in patterns of birth, maturation, aggression, cooperation, sex, and death. Even as populations of DNA strands evolve (keep that image in your mind clearly), they co-evolve with the environments they shape–and in turn, shape their fitness as replicating molecules. Thus, DNA is always evolving, but what we see evolving with DNA is a matter of scale at which we observe and investigate, whether we focus in on biochemical details of DNA function in cells, speciation and extinction, or the resilience or fragility of the whole biosphere that has shaped our evolution, even as we humans impact it.
REFERENCES
Dawkins, R. (1982a). The extended phenotype: the long reach of the gene. Oxford; New York: Oxford University Press.
Dawkins, R. (1982b). Replicators and Vehicles. In King’s College Sociobiology Group (Ed.), Current Problems in Sociobiology (pp. 45–64). New York: Cambridge University Press. Retrieved from http://www.stephenjaygould.org/library/dawkins_replicators.html
ELDREDGE, N., & Gould, S. J. (1972). Punctuated equilibria : an alternative to phyletic gradualism. In Schopf, Thomas J.M. (Ed.), Models in Paleobiology (pp. 82–115). San Francisco: Freeman, Cooper and Co.
Gingerich, P. D. (1983). Rates of Evolution: Effects of Time and Temporal Scaling. Science, 222(4620), 159–161. doi:10.2307/1691072
Gingerich, P. D. (1984). Punctuated Equilibria-Where is the Evidence? Systematic Zoology, 33(3), 335. doi:10.2307/2413079
Gingerich, P. D. (1985). Species in the Fossil Record: Concepts, Trends, and Transitions. Paleobiology, 11(1), 27–41. doi:10.2307/2400421
Gould, S. J., & Lewontin, R. C. (1979). The Spandrels of San Marco and the Panglossian Paradigm: A Critique of the Adaptationist Programme. Proceedings of the Royal Society of London. Series B. Biological Sciences, 205(1161), 581–598. doi:10.1098/rspb.1979.0086
Gould, Stephen Jay, & Eldredge, N. (1977). Punctuated Equilibria: The Tempo and Mode of Evolution Reconsidered. Paleobiology, 3(2), 115–151. doi:10.2307/2400177
Kimura, M. (1983). The Neutral Theory of Molecular Evolution. New York: Cambridge University Press.
Mayr, E. (1983). How to Carry Out the Adaptationist Program? The American Naturalist, 121(3), 324–334. doi:10.2307/2461153
Odling-Smee, F. J., Feldman, M. W., & Laland, K. N. (2003). Niche construction: the neglected process in evolution. Princeton: Princeton University Press.
Stuessy, T. F. (1987). Explicit Approaches for Evolutionary Classification. Systematic Botany, 12(2), 251–262. doi:10.2307/2419319
Vrba, E. S., & Eldredge, N. (1984). Individuals, Hierarchies and Processes: Towards a More Complete Evolutionary Theory. Paleobiology, 10(2), 146–171. doi:10.2307/2400395
Vrba, E. S., & Gould, S. J. (1986). The Hierarchical Expansion of Sorting and Selection: Sorting and Selection Cannot Be Equated. Paleobiology, 12(2), 217–228. doi:10.2307/2400492
